My Cart
0
You have no items in your shopping cart.
Creating an account has many benefits:

What is protein glycosylation?
One of the most common post-translational modifications of eukaryotic proteins is glycosylation. Glycosylation of proteins can affect many biological activities. For therapeutic glycoproteins, it can alter biological activity, targeting, transport, serum half-life, clearance and receptor recognition1,2. For this reason, biological manufacturers must monitor and characterize the glycosylation patterns of their recombinant therapeutic proteins3,4. There are two main glycosylation types of therapeutic proteins: N-linked polysaccharides and O-linked polysaccharides5. The attachment of N-polysaccharides begins in the endoplasmic reticulum (ER). In the endoplasmic reticulum, the core newborn polysaccharides are connected to the position with NXS/T sequence by side chain amide nitrogen on the specific asparagine (product number:F141104) of the protein, where X can be any amino acid residue except proline (product number:P111001). When the glycoprotein passes through the endoplasmic reticulum and Golgi apparatus, the N-sugar chain is pruned and further modified.Host cells and cell culture conditions can change the type of N-glycosylation on glycoproteins (from high mannose to complex and hybrid N-glycans). O-glycosylation occurs in the Golgi apparatus. N-glucan has a common core and consists of two N-acetylglucosamine (product number:N355747) (GlcNAc) residues and three mannose (Man) residues. But the only common core of O-glucan is the N-acetylgalactosamine (product number:N389642) (GalNAc) residue, which is connected by oxygen atoms on the side chain of the serine (product number:S329534
) or threonine residue of the protein.
Glycosidase: glycosylation is complex and heterogeneous, so a variety of analytical methods must be used to determine the structure of sugar chains and their position on glycoproteins. Glycosidase is an important tool and is usually used in conjunction with other analytical methods to remove, prune or modify polysaccharides. Glycosidase (glycoside hydrolase) (product number:G128643,N159659,A109181) is an enzyme that can break down the glycosidic bonds of complex sugars. These enzymes are used in three main areas of analysis of therapeutic glycoproteins: removing glucan for analysis, pruning glucan for sequencing, and modifying glucan in sugar engineering. Here, we describe the different enzymes used in each field and specific examples of their applications.
Extraction of polysaccharides for analysis
The most commonly used enzyme for the removal of N-sugar chains from glycoproteins is polypeptide-N-glycosidase F (PNGaseF) (product number:P420186). It is an amidase that cleaves between the innermost GlcNAc residue of the N-sugar chain and the asparagine residue, releasing the N-sugar chain, thus producing an aspartic acid (product number:A329587) residue instead of an asparagine residue on the protein. One of the main reasons why glycosidases are used so frequently is that they have a wide range of specificity and can cleave high-mannose, complex and mixed N-sugar chains. The only limitation of this enzyme is that it cannot be cut if α 1-3 fucose connects to the core GlcNAc residue (the sugar residue next to the protein). This modification occurs only in plants, insects, molluscs and parasitic worms. N-glucan containing this modification can be removed by PNGaseA enzyme. Once N-glucan is released by amidase, it can be flow-labeled for analysis by liquid chromatography-fluorescence detection (LC-FLD) or mass spectrometry (LC-MS) or capillary electrophoresis combined with laser-induced fluorescence detection (CE-LIF). Figure 1 illustrates the types of N-sugars released by PNGaseF from Erbitux therapeutic antibodies (cetuximab), which are detected by LC-MS analysis.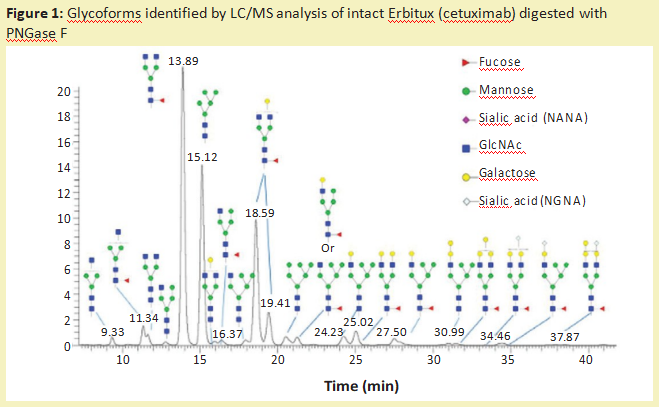
In addition to PNGaseF, many different endonuclease-β-N-acetyl glucosaminidases also release N-sugar chains from glycoproteins. Most endoglycosidases are limited to the type of N-sugar chain they can recognize and cleave. Table 1 shows the different properties of most commercially available Endo-β-N-acetylglucosaminidases. All of these endoglycosidases can hydrolyze the Nmagnetic N' diacetyl chitosan oligosaccharide part (located between the two GlcNAc residues on the N-glycosyl nucleus), resulting in the attachment of GlcNAc residues to the protein. Therefore, they are often used to determine the occupancy or presence of N-polysaccharides in specific sites. This is particularly useful for glycoproteins with multiple N-glycosylation sites.
Usually, the glycoprotein is digested with endoglucosidase and then cleaved with protease (product number:P298993). Then these peptide fragments were analyzed by MS. The extra mass of a single GlcNAc residue on any peptide fragment confirms that this position is occupied by the N-sugar chain6. Because the hydrolysis point of Endo-β-N-acetylglucosaminidase is far from the protein skeleton, these enzymes can usually cleave the N-sugar chain under non-denaturing conditions. This can happen even if these sites cannot cut PNGaseF under similar natural conditions. This is especially true for EndoS, which is specifically used for the hydrolysis of N-sugar chains in the Fc region of immunoglobulins and prefers non-denaturing conditions. This endoglycosidase has a particularly strong ability to hydrolyze sugar under natural conditions when the goal is to preserve the structure of the protein.
At present, there is no known broad-spectrum specific endoglycosidase that can separate all O-sugar chains from glycoproteins. Therefore, the analysis of O-sugar chain is much more difficult than that of N-sugar chain. There are two kinds of O-glycosidase available on the market. Streptococcus pneumoniae, which releases core 1 disaccharides, consists of galactose (Gal) and N-acetylgalactosamine (GalNAc) linked to β1-3, wherein the GalNAc residue is linked to the serine or threonine (product number:T108221,T100459) residue of the protein7. Another O-glycosidase comes from Enterococcus faecalis and has a wider specificity. It cleaves both core 1 disaccharide and core 3 disaccharide, in which core 3 disaccharide is connected to GalNAc by β1-38. If the disaccharides are further modified by sialic acid or other sugars, neither enzyme can be cleaved. The complete O-glucan can be chemically released by β-elimination method, but it must be noted that this method can not degrade the released glucan (product number:D140049,etc.).
Another major disadvantage of this method is that the protein is destroyed and its structure or activity can no longer be analyzed. Compared with the complete chemical release of O-glucan, a combination of exoglycosidase and O-glucosidase can be used to prune O-glucan (figure 2). Although this method can retain the structure and activity of the protein, it will degrade the polysaccharide, so it can not be characterized. In addition, some O-glucans contain carbohydrate modifications (e.g., vulcanization or acetylation) so that the modified sugars are not cut by exoglycosidase.
Prune the sugar chain for sequencing
Extracellular glycosidase is an important enzyme that removes polysaccharides from the non-reducing ends of polysaccharides one at a time. Commercially, there are a large number of different exoglycosidases. Extracellular glycosidase is specific to the types of sugars and their isomers (α or β). Some exoglycosidases are common and can break down many different connections. For example, broad-specific neuraminidase (product number:N128387) can remove sialic acid residues from α2-3, α2-6, α2-8 or α2-9 linked to sugar chains. Other exoglycosidases are more specific for specific linkage. For example, β1-4 galactosidase can only remove β1-4 linked galactose. Because of the innate specificity of these enzymes, exoglycosidase is useful for pruning and sequencing sugar chains. Basic LC-MS or matrix-assisted laser desorption/ionization (MALDI) analysis can only determine the size of polysaccharides. However, in order to determine the connection or type of monosaccharides present on polysaccharides, it is necessary to perform tandem mass spectrometry (MS-MS), such as collision induced dissociation (CID) or using exoglycosidase, followed by CE or MS analysis.
When using exoglycosidase, it is important to consider the quality of the enzyme preparation. Enzyme preparations isolated from natural sources are often contaminated by other glycosidase activities, which makes it more difficult to elucidate the structure of polysaccharides. The most commonly used recombinant sequencing version of exoglycosidase can now be used commercially for sensitive dextran analysis workflows.
Figure 3 shows how glycosidase sequenced the polysaccharides released from the Enbrel therapeutic protein (enalapril). Extracellular glycosidase can also be used to sequence sugar chains that are still attached to glycoproteins. These enzymes are particularly useful for analyzing and detecting potential antigenic structures on sugar chains. For example, treating therapeutic proteins with universal alpha-galactosidases, such as those from green coffee beans, can help identify low levels of galactose α 1-3Gal (α-galactose epitopes) that are not present in humans and can produce immunogenicity 9.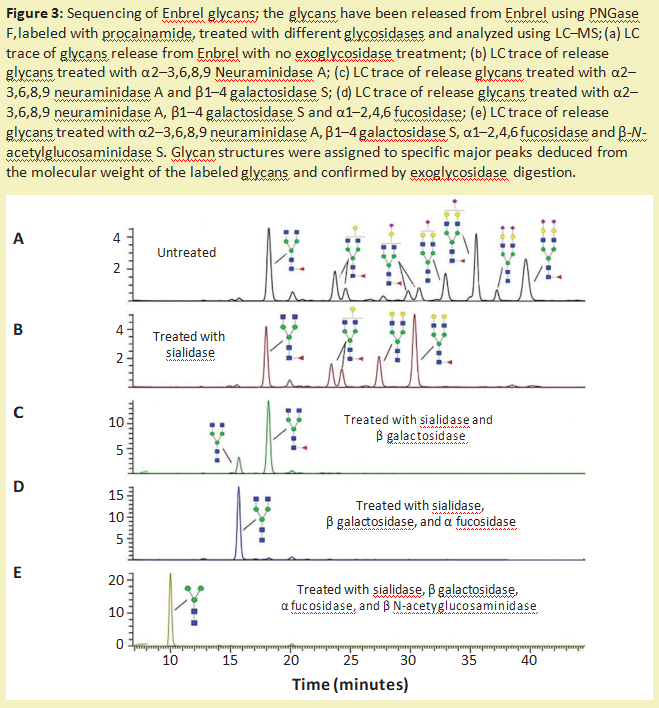
Modification of glucan
In addition to removing the complete sugar chain and sequencing the sugar chain, glycosidase can also be used to modify the sugar chain on glycoprotein10 (called sugar engineering). One way is to use endoglucosidase to remove unwanted N-linked polysaccharides. Then, through the GlcNAc residue left by the cleavage of endoglucosidase, the uniform glucan produced by chemical enzyme synthesis can be connected to the glycoprotein. Endoglucosidase (naturally transglycosylation), such as Endo-M11, or an endoglucanase mutant that drives the reaction towards sugar addition rather than sugar removal can be used to achieve the desired glucan ligation12.(figure 4)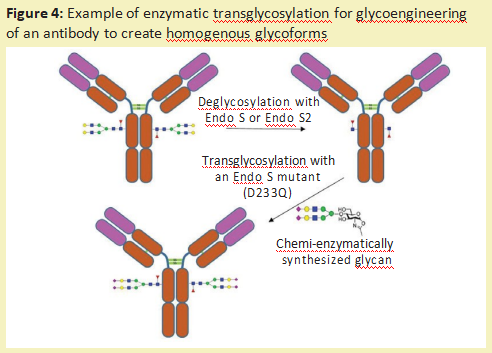
Another method of sugar engineering is to use exoglycosidase to trim the sugar chain on the glycoprotein to a uniform size. The polysaccharide can then be reconstructed with a specific glycosyltransferase (product number:G293642), which transfers monosaccharides from the ribonucleic acid donor to the sugar chain. This produces a glycoprotein with a more uniform sugar chain structure (figure 5).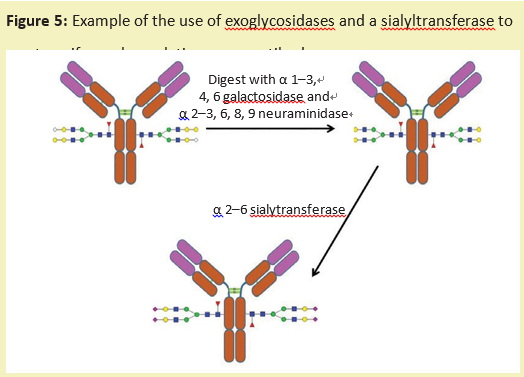
Future applications
So far, most of these in vitro sugar engineering methods are used on a small scale. The exception is glycoprotease, which is used to treat lysosome storage diseases. Several of these enzymes have been used by sugar engineering to expose the mannose structure of proteins through receptor-mediated endocytosis to improve enzyme transport to lysosomes13. In addition, the modification of glycosylation pathways in host cells is being used to produce therapeutic proteins with the required glycosylation. The production of therapeutic proteins in these cell lines is well under way, and several therapeutic proteins are in clinical trials. Scientists predict that as the demand for high-throughput methods increases, the speed and simplicity of dextran analysis will improve with further improvements in enzymes and analysis. New antibody-specific proteolytic enzymes, such as IDES and IDEZ, are improvements on typical antibody trypsin / lysozyme C or papain (product number:P128675,P128674) to digest antibodies because they specifically cleave in the hinge region of the antibody rather than at the secondary site. Using these enzymes and improved software, the constant region of antibodies can be analyzed by MS. This makes it possible to determine the type of polysaccharides present on the FC without the need to remove, purify and label N-polysaccharides. Other improvements may lead to the discovery of new enzyme specificity. For example, an enzyme that is widely specific to complex O-glucans can better analyze these types of protein modifications. As more therapeutic proteins are expressed in transgenic plant and insect cell lines, PNGase that removes all N-sugars will be useful. Finally, as dextran analysis methods become more refined, higher throughput and higher sensitivity, it should be possible to choose cloning or use a selected sugar engineering expression host to produce antibodies with the desired glycosylation pattern during the development process. This speed and sensitivity will also allow for continuous monitoring of the fermentation process, which will increase the production of therapeutic glycoproteins.
References
1. Ohtsubo K,Marth JD.Glycosylation in Cellular Mechanisms of Health and Disease.Cell 126(5)2006:855–867;doi:10.1016/j.cell.2006.08.019.
2. Walsh G.Posttranslational Modifications of Protein Biopharmaceuticals.Drug Discovery Today 15(17–18),2010:773–780;doi:10.1016/j.drudis.2010.06.009.
3. Beck A,et al.Trends in Glycosylation,Glycoanalysis,and Glycoengineering of Therapeutic Antibodies and Fc-Fusion Proteins.Curr.Pharm.Biotechnol.9(6),2008:482–501.
4. Sethuraman N,Stadheim TA.Challenges in Therapeutic Glycoprotein Production.Curr.Op.Biotech.17(4)2006:341–346;doi:10.1016/j.copbio.2006.06.010.
5. Spiro RG.Protein Glycosylation:Nature,Distribution,Enzymatic Formation,and Disease Implications of Glycopeptide Bonds.Glycobiology 12(4)2002:43R–56R.
6. Wang L,et al.Structural Analysis of a Highly Glycosylated and Unliganded gp120-Based Antigen Using Mass Spectrometry.Biochem.49,2010:9032–9045;doi:10.1021/bi1011332.
7. Fujita K,et al.Identification and Molecular Cloning of a Novel Glycoside Hydrolase Family of Core 1 Type O-Glycan-Specific Endo-Alpha-N-Acetylgalacto-saminidase from Bifidobacterium longum.J.Biol.Chem.280(45)2005:37415–37422;doi:10.1074/jbc.M506874200.
8. Koutsioulis D,Landry D,Guthrie EP.Novel Endo-N-Acetylgalactosaminidases with Broader Substrate Specificity.Glycobiol.18(10).2008:799–805;doi:10.1093/glycob/cwn069.
9. Bosques CJ,et al.Chinese Hamster Ovary Cells Can Produce Galactose--1,3-Galactose Antigens on Proteins.Nat.Biotechnol.28(11)2011:1153–1156;doi:10.1038/nbt1110-1153.
10. Rich JR,Withers SG.Emerging Methods for the Production of Homogeneous Human Glycoproteins.Nat.Chem.Biol.5(4)2009:206–215;doi:10.1038/nchembio.148.
11. Yamamoto K,et al.Transglycosylation Activity of Mucor hiemalis Endo-Beta-N-Acetyl-Glucosaminidase Which Transfers Complex Oligosaccharides to the N-acetylglucosamine Moieties of Peptides.Biochem.Biophys.Res.Comm.203(1)1994:244–252.
12. Huang W,et al.Chemoenzymatic Glycoengineering of Intact IgG Antibodies for Gain of Functions.J.Am.Chem.Soc.134(29)2012:12308–12318;doi:10.1021/ja3051266.
13. SoláRJ,Griebenow K.Glycosylation of Therapeutic Proteins.BioDrugs.24(1)2010:9–21;doi:10.2165/11530550-000000000-00000.
14. Trimble RB,Tarentino AL.Identification of Distinct Endoglycosidase(Endo)Activities in Flavobacterium meningosepticum:Endo F1,Endo F2,and Endo F3.Endo F1 and Endo H Hydrolyze Only High Mannose and Hybrid Glycans.J.Biol.Chem.266(3)1991:1646–1651.
15. Tarentino AL,et al.Multiple Endoglycosidase F Activities Expressed By Flavobacterium meningosepticum Endoglycosidases F2 and F3:Molecular Cloning,Primary Sequence,and Enzyme Expression.J.Biol.Chem.268(13)1993:9702–9708.
16. Collin M,Olsén A.Endo S:A Novel Secreted Protein from Streptococcus pyogenes with Endoglycosidase Activity on Human IgG.EMBO J.20(12)2001:3046–3055.
17. Sjögren J,et al.EndoS 2 Is a Unique and Conserved Enzyme of Serotype M49 Group A Streptococcus That Hydrolyses N-Linked Glycans on IgG and1-Acid Glycoprotein.Biochem.J.455(1)2013:107–118;doi:10.1093/emboj/20.12.3046.
18. Mizuochi T,Amano J,Kobata A.New Evidence to the Substrate Specificity of Endo-Beta-N-Acetlylglucosiamindase D.J.Biochem.95(4)2008:1209–1213.
19. Yamamoto K,et al.Novel Specificities of Mucor hiemalis Endo-Beta-N-Acetylglucosaminidase Acting Complex Asparagine-Linked Oligosaccharides.Biosci.Biotechnol.Biochem.58(1)1994:72–77.
List of related products