My Cart
0
You have no items in your shopping cart.
Creating an account has many benefits:

The TGF-β family (Transforming growth factor-beta family) includes members of TGF-β1, TGF-β2, TGF-β3, BMP (Bone morphogenetic protein), Activin, and Nodal, which play important roles in a variety of biological processes including cell proliferation, differentiation, apoptosis, extracellular matrix synthesis, and immune regulation. The responsiveness of the TGF-β family can be regulated by proteases targeting the TGF-β receptor and co-receptors. The hydrolytic activity of these proteins increases the regulatory complexity of TGF-β family signaling. For example, thrombin cleavage of PAR1 leads to the internalization of co-receptor endothelial glycoproteins, as well as MMP-14 and MMP-16-mediated shedding of biglycan [1-2]. The BMP-1 protease family also play an important role in regulating TGF-β family activity. The highly conserved BMP-1/PCP subgroup of the astaxanthin family includes BMP-1, selectively spliced mammalian toxoid (mTLD), mammalian toxoid 1, and mTLL2. These enzymes contain an astaxanthin protease structural domain, CUB, and an EGF-like structural domain. They cleave the N-terminus to aspartic acid residues in a variety of extracellular matrix (ECM) components (collagen, laminin, small leucine-rich proteoglycans, and SIBLING proteins), lysyl oxidases, and related molecules such as growth factors [3].
TGF-β family proproteins are cleaved in the trans-Golgi between the N-terminal propeptide and mature growth factors. For TGF-β1, -β2, -β3, GDF-8, and GDF-11, the pre-structural domain binds to growth factors for secretion and maintains growth factors in an inactive state. The BMP-1 protease family regulate the activation of these potential complexes by several mechanisms.
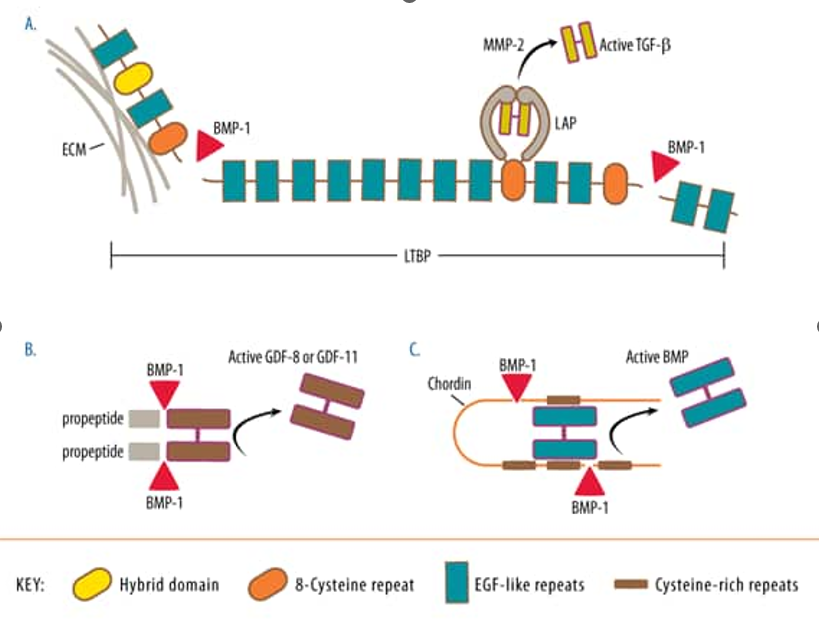
Large potential complexes consisting of TGF-β, potential associated peptide (LAP) and potential TGF-β binding protein (LTBP) bind to the ECM via LTBP. A recent report by Ge et al [4]. described how BMP-1 cleaves LTBP1 at two sites, leaving the central portion associated with TGF-β/LAP and severing the complex from the ECM, as shown in Fig. 1A. The processes associated with LTBP1 are required for MMP-2-mediated efficient release of TGF-β from LAP. Knockout mice lacking BMP-1, mTLD, and mTLL1 greatly increased the number of potential complexes associated with the ECM and significantly reduced the levels of active TGF-β. One of the many roles of TGF-β is to induce further expression of BMP-1, which leads to positive feedback regulation of TGF-β activity. Certain TGF-β family members retain noncovalent binding to propeptide after cleavage from the underlying protein. For example, BMP-1 family proteases cleave at a single site of non-covalently bound GDF-8 and GDF-11 protein fragments to release active growth factors, as shown in Fig. 1B [5-6]. BMP-1, mTLL-1, and mTLL-2 are comparable in their effects on this activity[3]. The Asp-substituted GDF-11 prodomain in the cleavage site binds to mature GDF-11 and blocks its activity [6].
Other BMP homodimers and heterodimers are not secreted in complex with their precursors, but remain potentially complexed by binding to chordin. Activation of these complexes is achieved by BMP-1-mediated proteolytic hydrolysis of two sites within Chordin, as shown in Figure 1C [7]. Recognition of chordin is conferred by the first CUB structural domain of BMP-1 because mTLL-2 does not cleave chordin unless its first CUB structural domain is interchanged with that of BMP-1. [8-9]
BMP-1 family protease activity is required for embryonic development. Knockouts result in fatal defects in embryonic or perinatal skull, heart, and abdominal wall formation. In these experiments, there is reduced TGF-beta release from large latent complexes, inefficient chordin processing, and abnormal collagen fiber production. [10-11]
References
1. Tang, H. et al. (2005) Blood 105:1977.
2. Velasco-Loyden, G. et al. (2004) J. Biol. Chem. 279:7721.
3. Ge, G. & D.S. Greenspan (2006) Birth Defects Res. 78:47.
4. Ge, G. & D.S. Greenspan (2006) J. Cell Biol. 175:111.
5. Wolfman, N.M. et al. (2003) Proc. Natl. Acad. Sci. USA 100:15842.
6. Ge, G. et al. (2005) Mol. Cell. Biol. 25:5846.
7. Scott, I.C. et al. (2001) Nature 410:475.
8. Petropoulou, V. et al. (2005) J. Biol. Chem. 280:22616.
9. Moali C. et al. (2005) J. Biol. Chem. 280:24188.
10. Pappano, W.N. et al. (2003) Mol. Cell. Biol. 23:4428.
11. Suzuki, N. et al. (1996) Development 122:3587
List of related products